
Introduction
Soil fumigants are pesticides that, when applied to the soil, form a gas to control pests that disrupt plant growth and crop production. Fumigation suppresses weeds and soilborne plant pathogens in crops such as potatoes, onions, sugar beets, tomatoes, mint, carrots and strawberries in the Pacific Northwest.
Chemical fumigation is widely practiced. Globally, agricultural producers applied 2.17 million tons of pesticides (active ingredients) to crops in 2020. U.S. producers applied 0.37 million tons. Common chemical fumigants used in the United States are chloropicrin, 1,3-dichloropropene, metam sodium and dazomet. While chemical fumigation has been shown to be effective, it is expensive. Recent research also shows that it impacts soil health.
There is another way for producers to use fumigants that does not involve synthetic chemicals. A practice known as biofumigation is an eco-friendly alternative that employs cover crops such as mustard, canola and oilseed radish. In this publication, producers will learn how these crops, when incorporated into the soil at the right time and in the correct manner, preserve and enhance soil health while combating pests, diseases and weeds.
Threats to production
Pests and weed pressure significantly impact yield. In 2018, pests such as weeds, pink root disease, thrips and plant-parasitic nematodes reduced onion yield in the Treasure Valley by 14%. Yellow nutsedge is the major weed problem in onions. A 2022 Oregon State University survey found pink root was the most significant plant pathogen in Treasure Valley dry bulb onions. Pink root infected 64% of surveyed acreage and reduced yields by 1.8%.
In potatoes, the potato early dying disease complex reduced yield by 30% to 50%. Verticillium spp. is the primary pathogen of PED, and the presence of root-lesion nematodes (Pratylenchus spp.) exacerbates disease severity and yield loss.
Nationally, the harvested acreage for onion and potato in 2020 was 134,700 and 911,700 acres, respectively. The Pacific Northwest and California are among the top producers of potatoes, sugar beets and onions.
|
Crop |
Ranking |
Acres |
|---|---|---|
|
Potatoes |
Fourth | 45,000 |
|
Strawberries |
Third | 1,100 |
|
Sugar beets |
10th | 19,800 |
Source: Oregon Agriculture Statistics and Directory, 2022
Pros and cons of chemical fumigants
Studies have found that chemical fumigation works. It was shown to be the most effective approach to controlling weeds, pathogens and soilborne diseases in the Treasure Valley of Eastern Oregon and southwestern Idaho. Fumigation controls weeds such as yellow nutsedge (Cyperus esculentus) and the pathogens responsible for pink root and potato early dying.
While it has proven successful, chemical fumigation is expensive. At a cost of approximately $741 per hectare, a 2024 study found the cost of fumigating potato acreage would add up to about $13.5 million in Oregon alone.
Besides economic costs, a 2010 study found that repeated application of these chemical fumigants damaged soil health by reducing microbial biomass and enzyme activities; altering the structure of microbial communities; and affecting carbon/nitrogen cycling and mineralization.
A 2021 study found that chemical fumigation alters soil carbon transformation. When comparing soils treated with fresh barley plant residues to those treated with metam sodium, researchers observed lower respiration rates (428 mg CO2 C kg-1 dry soil) and a higher percentage (22%) of leftover plant residues in the metam sodium-fumigated soil.
In addition to the carbon cycle, studies have reported that chemical fumigation with metam sodium or chloropicrin affects the soil nitrogen cycle by significantly inhibiting nitrification. Chemical fumigants can kill soil microorganisms through direct contact, albeit in a nonselective manner, particularly when used at high concentrations.
Fumigation and soil health
Agricultural practices can positively or negatively impact soil health by modifying the physical, chemical and biological processes that control nutrient mineralization, soil aggregate formation, soil mineral sorption, microbial processing of residue and carbon transport into the subsoil.
Soil health is a crucial aspect of sustainable agriculture, as it directly influences crop productivity and environmental sustainability. Soil health depends on the maintenance of four major functions: carbon transformations, nutrient cycles, soil structure maintenance, and the regulation of pests and diseases.
Soil health is essential to maintaining fertility, structure and nutrient cycling. Potential alternatives to chemical fumigation, such as biofumigation cover crops and green manure, are needed to manage soilborne diseases, enhance soil health, increase microbial biodiversity and improve productivity.
There are many types of cover crops, from legumes to grasses and brassicas. Each benefits soil health in unique ways.

What is biofumigation?
Biofumigation is the process of incorporating glucosinolate-containing crops such as Brassicas to suppress soilborne pests. Glucosinolates are diverse secondary metabolites found in various Brassica species, such as mustard, oilseed radish, arugula (Figure 1) and related plants, playing a crucial role in plant defense mechanisms.
The types and concentrations of glucosinolate can vary significantly among different species and even among cultivars within the same species. These complex sulfur-containing compounds are stored in the vacuoles of plant cells, separate from the enzyme myrosinase, which catalyzes glucosinolate breakdown. This compartmentalization prevents the premature release of bioactive products.
When tissue of these plants is damaged by chopping, tilling or other mechanical disruption, glucosinolate and myrosinase come into contact. This initiates a chemical reaction in the presence of moisture that releases isothiocyanates, potent biofumigants that are toxic to pathogens and pests. ITCs are chemically similar to the active ingredient in the commonly used chemical fumigantVapam® (Natriun N-methyl-dithiocarbamate). The enzymatic activity of myrosinase is not limited to plant tissues; it can also be found in certain soil fungi and microbial communities, further contributing to glucosinolate hydrolysis when cover crop residues are incorporated into the soil.
Biofumigation cover crops and agronomic management
The plant families Brassicaceae, Caricaceae and Capparaceae contain more glucosinolate than other plants. Brassicaceae produce high levels of glucosinolate (Table 2) that are similar to the compounds released from the commercial chemical fumigant metam sodium (methyl isothiocyanate). Brassicas have shown potential for managing soilborne pests, including Verticillium dahliae and plant-parasitic nematodes.
Biofumigation cover crops not only suppress soilborne pathogens and pests but also attract pollinators with their abundant, nectar-rich, vibrantly colored flowers (Figure 2). These blooms provide a critical source of nectar and pollen, especially when other floral resources are scarce, supporting the health and diversity of bees, butterflies and other pollinators. Additionally, the habitat enhancement created by these crops fosters biodiversity and ecosystem stability, offering shelter for beneficial insects while potentially boosting yields of nearby pollinator-dependent crops.
Follow crop recommendation guidelines from seed suppliers for seeding rates, planting time, fertilization and irrigation to maximize the effectiveness of biofumigation cover crops. Proper seeding rates ensure adequate biomass production. Plant at the right time to optimize growth and glucosinolate levels. Apply fertilizers according to recommended guidelines to enhance biomass and biofumigation potential. Maintain adequate soil moisture through recommended irrigation practices to support crop establishment and growth.

|
Common name |
Scientific name |
Characteristics |
Glucosinolate content (µmol/g DW) |
References |
|---|---|---|---|---|
|
Salad rocket |
Eruca sativa | Fast-growing annual with high glucosinolate content; effective for weed suppression; can be used as a leafy vegetable. | 11–125 | Kim and Ishii (2006) |
|
Black mustard |
Brassica nigra | High glucosinolate content; strong biofumigant properties; suitable for suppressing nematodes and certain soilborne pathogens. | 80–120 | Li et al. (2021) |
|
Indian mustard |
Brassica juncea | Contains high levels of glucosinolates; effective biofumigant; vigorous growth; tolerates a wide range of soils; drought-resistant. | 53–140 | Huang et al. (2022) |
|
Yellow mustard |
Sinapis alba | Lower glucosinolate content compared to other mustards; quick growth; good for weed suppression but less effective for pathogen control. | 10–30 | Cámara-Martos et al. (2021) |
|
Canola/rapeseed |
Brassica napus | Moderate glucosinolate levels; excellent biomass producer; provides good soil cover. Commonly used as a rotational crop to improve soil health. | 30–60 | Jhingan et al. (2023) |
|
Cole crops |
Brassica oleracea | Includes cabbage, broccoli, and kale; moderate glucosinolate content; effective in breaking disease cycles but may require careful management. | 4–24 | (Bhandari et al., 2020) |
|
Turnip |
Brassica rapa | Quick-maturing cover crop; moderate glucosinolate levels; roots and leaves decompose to release biofumigant compounds; suitable for erosion control | 4–54 | Kim et al. (2023) |
|
White mustard |
Sinapis alba | High biomass production; relatively lower glucosinolate content; useful for erosion control and organic matter addition but less effective as a biofumigant. | 6–54 | Petersen et al. (2018) |
|
Oilseed radish |
Raphanus sativus | Deep taproot; breaks up soil compaction; moderate glucosinolate levels; decomposes quickly to release biofumigant compounds; excellent for soil aeration. | 20–50 | Tsytsiura (2024) |
Cover-crop termination
The timing and method of cover crop termination are critical to biofumigation’s success. Manage these factors to release natural compounds into the soil, target pests and pathogens, and improve soil conditions for subsequent crops.
Termination timing
Effective pest control strategies depend heavily on the termination timing of crop growth stage and method. Terminating biofumigation crops like mustard or radish at the flowering stage (Figures 2 and 3) or just before seed set is crucial because it maximizes the concentration of glucosinolates within the plant. These are the key compounds responsible for producing the volatile, pest-suppressing ITCs when the plant tissue is chopped and incorporated into the soil, thus achieving the most effective biofumigation action against soilborne pathogens and pests.
Research shows that terminating mustard at early flowering can result in up to 40%–60% higher pest suppression compared to termination at later stages. One study also highlighted that glucosinolate concentrations vary across plant developmental stages, with levels peaking at flowering, and noted that root glucosinolate concentrations are highest at early growth stages and diminish as the plant matures. To harness the maximum biofumigant potential, carefully time termination — preferably at the flowering stage — to ensure optimal glucosinolate availability.

Techniques for terminating and incorporating biofumigants
Mechanical maceration techniques such as flail-mowing, roller-crimping or rotary tilling are highly effective at maximizing glucosinolate hydrolysis (Figure 4). Biofumigant cover crops should be finely chopped into smaller fragments to increase the surface area for enzyme-substrate interactions. The more thoroughly the plant tissues are macerated, the more complete the enzymatic conversion of glucosinolate to ITCs. This process accelerates the biofumigant action and enhances the overall effectiveness of pest and pathogen suppression.
The residues should be incorporated into the soil immediately using equipment like a rototiller. Research in 2022 demonstrated that plant biomass should be incorporated into the soil immediately to preserve ITC concentrations. A delay of just four days can significantly reduce its effectiveness.
Field trials on oilseed radish and brown mustard in 2020 examined the effects of different termination methods. The study found that combining mechanical tissue maceration, soil incorporation with a handheld rototiller and the use of plastic covers significantly enhanced the glucosinolate-to-ITC conversion process. Covering the soil with black plastic sheeting after incorporation helps to trap volatile ITCs, extending their fumigation effect and improving pest and pathogen control. Covering soil with plastic mulch also creates a microenvironment that retains moisture and raises soil temperatures, further boosting the biofumigation process, especially under high tunnels.
In large-scale operations, cultipacking or other sealing methods, such as irrigation and plastic tarps, are recommended to trap the biofumigation gases in the soil. Overall, for biofumigant crops to reach their full potential, growers must carefully manage termination timing and incorporate strategies like irrigation and soil coverage to optimize ITC release and efficacy.


Optimal condition for release of biofumigant compounds
The hydrolysis process of glucosinolate into ITCs depends on environmental conditions, including soil moisture and temperature. Rapid soil incorporation of biofumigant crops immediately after termination is essential to minimize the volatilization and loss of ITCs to the atmosphere. A 2009 study emphasized the importance of soil moisture, noting that irrigating the soil before or right after incorporation enhances ITC concentrations. Moisture promotes myrosinase activity and improves the conversion efficiency of glucosinolate to ITCs, making the biofumigant effect more pronounced. This highlights the necessity for integrated management practices that consider soil conditions alongside crop termination.
Impact of biofumigant cover crops
Plant diseases
Soilborne diseases are a significant challenge to crop production, leading to reduced crop performance, decreased yields and increased production costs worldwide. This section covers the control of soilborne pathogens using biofumigant cover crops.
Research in France highlighted that planting Brassica juncea reduced the incidence of root rot (caused by Rhizoctonia solani) in sugar beets. The severity of the disease and R. solani incidence level reduced in subsequent years after incorporating mustard cover crops in soil from nearly 30% in 2005 to 15% in 2007, which is significantly lower than the control bare soil.
In-vitro investigations on the impact of Brassica species on Phytophthora spp. demonstrated that B. juncea inhibited the mycelial growth of Phytophthora spp.
Rotating crops with canola and rapeseed (B. napus) helps reduce Rhizoctonia disease in potatoes.
Biofumigation cover crops can also help manage soilborne diseases in nursery soils. In 2020 greenhouse trials, yellow mustard (White Gold), arugula (Astro), mighty mustard (Pacific Gold), rape (Dwarf Essex), turnip (Purple Top Forage), brown mustard (Kodiak) and mustard green (Amara) effectively reduced disease severity from R. solani and P. nicotiane in viburnum and hydrangea, except for radish, when incorporated two weeks before planting.
Brassica crops like B. nigra, B. juncea and B. carinata release high amounts of propenyl isothiocyanate, which helps suppress Fusarium oxysporum in nursery soils.
Using brassica cover crops can be an effective strategy for disease management through biofumigation.
Nematode control
Nematodes are microscopic worms present in almost all environments, from aquatic to terrestrial (soil). The majority of nematode species play crucial roles in nutrient cycling and organic matter decomposition. But some are plant-parasitic and pose significant threats to agricultural production. Species such as root-knot nematodes (Meloidogyne spp.), cyst nematodes (Heterodera spp.) and root-lesion nematodes (Pratylenchus spp.) feed on plant roots, causing damage that leads to reduced water and nutrient uptake, stunted growth and yield losses.
Chemical fumigation and synthetic nematicides are commonly used to control nematodes, which can be expensive and environmentally harmful. As a result, alternative strategies, such as the use of biofumigant cover crops, are gaining popularity as eco-friendly and cost-effective solutions for nematode management.
A greenhouse study in New Mexico showed that broccoli (Arcadia) had the least root-knot nematode damage, while mustard (Caliente 61, Caliente 199 and Pacific Gold) increased nematode numbers. In 2012, yellow mustard was found to decrease population densities of the plant-parasitic nematode Globodera rostochiensis in potatoes. A greenhouse study in Australia found that mixing broccoli tissue into nematode-infested soil reduced root-knot nematode (M. incognita) levels, with the highest rate (20 g per pot) lowering infestation to 5%, compared to 100% in untreated soil.
Brassica cover crops can suppress root-knot nematodes, but effectiveness varies by species, location and biomass produced. In Australia, fodder radish significantly reduced root-knot nematodes in vegetables, showing resistance due to genetic traits rather than just glucosinolates. Another study found that cultivating radish as a cover crop effectively reduced the egg population of M. japonica, supporting their use in nematode management.
Soil health
Soil is a viable living ecosystem that helps sustain the lives of plants, humans and animals. Soil has many functions. Some of its key roles include maintaining the soil microbiome, filtering heavy metals and pollutants, infiltrating water and cycling nutrients.
Soil health can be maintained by maximizing soil cover, biodiversity and living roots while reducing physical disturbance and agrochemical use. Besides controlling diseases and applying nematode pressure, biofumigant cover crops maintain soil health by:
- Increasing nutrient cycling.
- Retaining water.
- Stabilizing aggregate.
- Contributing organic matter.
- Reducing groundwater pollution by preventing nitrate leaching.
Studies have demonstrated the potential of cover crops to enhance soil health and nutrient dynamics across different cropping systems. A 2022 study evaluated the performance of brassica cover crops — brown mustard, fodder radish and turnip rape — alongside the legume purple vetch in a sunflower rotation. Findings indicated that fodder radish produced the highest biomass, while brassicas exhibited a significantly higher carbon-to-nitrogen (C:N) ratio than legumes. Preceding sunflower planting, brown mustard plots had the greatest nitrogen availability (200 lb ha-1), and both fodder radish and purple vetch contributed to increased soil mineral nitrogen.
In a chile pepper rotation in southern New Mexico, researchers investigated the biofumigant potential of mustard (Caliente 61, Caliente 199 and Pacific Gold) and broccoli (Arcadia). In the first year, Pacific Gold produced substantial biomass (1,241 kg ha-1), leading to increased soil organic matter (1.12%), reduced soil pH (7.44), and improved water infiltration compared to fallow plots. However, in the second year, poor biomass production resulted in no significant increase in soil organic matter. Similarly, another study assessed the benefits of white mustard as a cover crop in potato-wheat rotations and found that its inclusion reduced the incidence of Verticillium dahliae while enhancing soil water infiltration.
Research in the Pacific Northwest investigated the effects of white mustard incorporation on microbial biomass carbon across various soil types, including Quincy sand, Quincy loamy fine sand, Timmerman sandy loam, Warden silt loam and Shano silt loam. Results indicated that white mustard significantly increased microbial biomass carbon in all soil types except Warden silt loam. The highest microbial biomass carbon was observed in Shano silt loam, with approximately 260 mg C/kg of soil after mustard incorporation, outperforming chemical fumigation (220 mg C/kg).
A 2009 study found that incorporating B. juncea residues increased porosity in the topsoil (0–10 cm) compared to bare soil. Similarly, a study in Tasmania comparing ryegrass, brassicas and fallow treatments found that cover crops significantly enhanced soil microbial communities. Both brassicas and ryegrass increased bacterial, fungal and eukaryotic populations, with B. juncea showing particularly strong effects two and four weeks post-incorporation compared to fallow plots.

Weeds
In agricultural, forestry and rangeland systems, as well as in natural landscapes, it is often essential to reduce the presence of weeds to meet land-use objectives. Weeds compete with crops for nutrients, water, space and light. They also serve as alternative hosts for disease-causing microorganisms and pests. Weeds are widely controlled with herbicides. Herbicides initially worked well, but more recently, researchers identified weeds resistant to almost 21 herbicide modes of action. Currently, 530 weeds are known to be resistant to herbicides, according to the International Herbicide-resistant Weed Database. Nearly 50% of them are found in the U.S. and Australia.
In addition to resistance problems, herbicides could harm the environment through drift and hazardous leaching into groundwater.
As a countermeasure to herbicide resistance, focus has shifted to controlling weeds with biological measures — the most common being cover crops. Biofumigant cover crops have started to attract researchers due to the release of allelochemical ITC, which has the potential to inhibit weed seed germination (Figure 5).
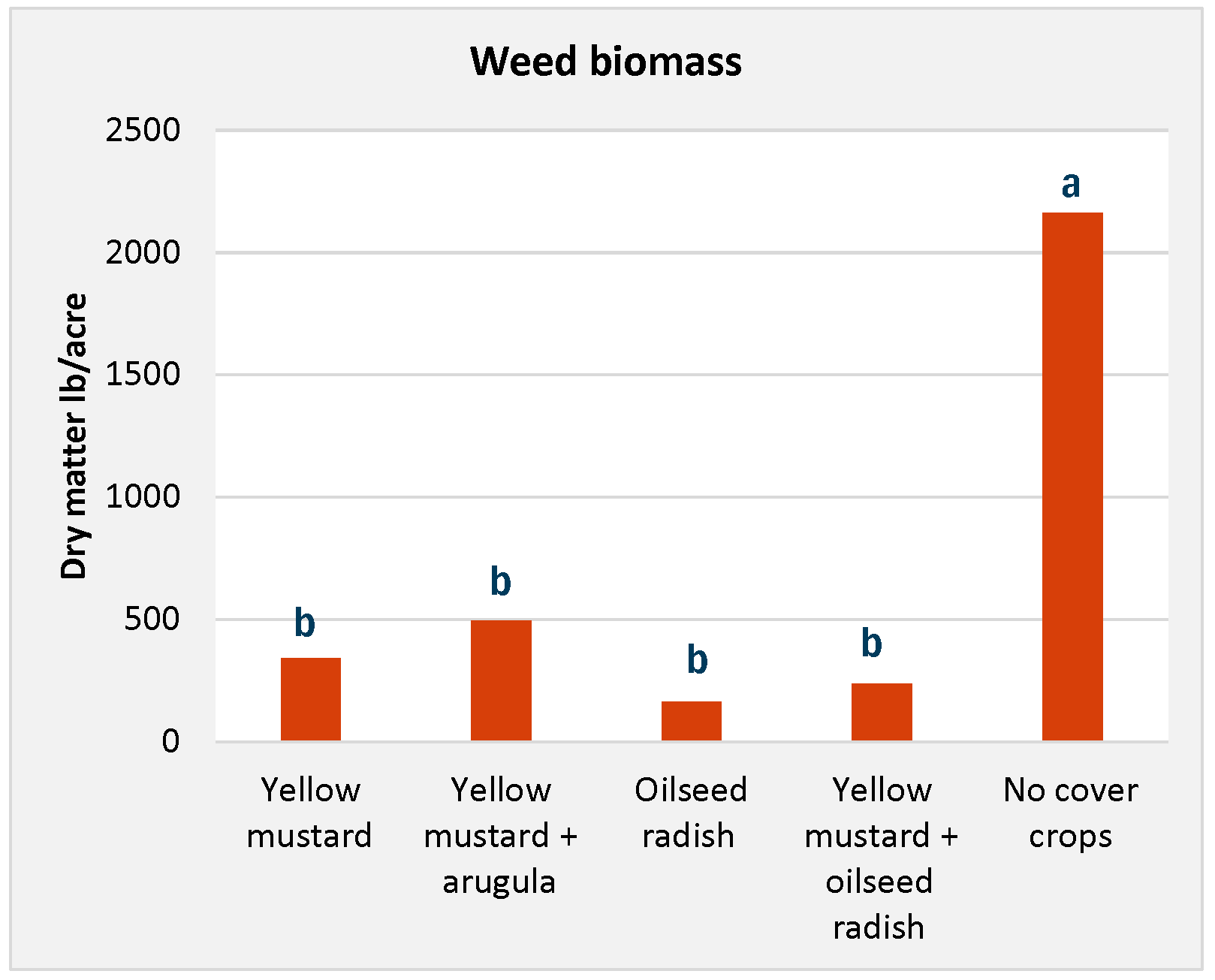
A study in Canada examined the impact of Indian mustard and oat cover crops on weed suppression. The treatments included different mustard-oat combinations planted in spring and fall. Initially, Indian mustard exhibited limited weed-suppression capabilities. But by 2015 and 2016, its effectiveness increased, likely due to higher glucosinolate production from potassium sulfate application. In spring 2015, mustard-based treatments significantly reduced weed density from about 2,200 to more than 600 plants per square meter. Similar reductions were observed in fall 2015 and 2016 compared to the control (no cover crops).
A 2009 study found that brown and yellow mustard cover crops reduced hairy galinsoga (Galinsoga quadriradiata) biomass by 90%–99% over two years. However, other studies suggest that brassicas may suppress weeds by immobilizing nitrogen, which can negatively impact certain crops like brome grass and barley. Additionally, brassica biofumigation has been reported to reduce oilseed crop yield.
These variations in weed suppression and crop impact likely depend on soil conditions, glucosinolate levels, and the timing and method of incorporation.

Summary and future interest
Biofumigation is a growing alternative to chemical fumigation. It is effective in controlling nematodes, diseases and weeds. Additionally, biofumigation cover crops help improve soil health.
The effectiveness of biofumigant crops in agricultural systems depends on a holistic approach that integrates proper crop selection, strategic timing and optimized termination methods. Managing environmental factors such as soil moisture and temperature, in combination with mechanical practices, ensures maximum ITC production and persistence.
Ultimately, a well-planned biofumigation strategy can significantly contribute to sustainable integrated pest and disease management, reduce the reliance on chemical fumigants, and improve soil health in the long term.
Resources
- Mustards | Sustainable Agriculture Research & Education Program, University of California, Davis
- Canola/Rape | Sustainable Agriculture Research & Education Program, University of California, Davis
- Cover Crop Fact Sheet Forage Turnip and Rape, Cornell University
- Radish as a cover crop | Integrated Crop Management, Iowa State University
- Torabian, S., E. Kim, R. Qin, V. Sathuvalli, H.T. Gollany, M. Kleber. 2025. Soil microbial biomass influenced by cover crop after fumigation of potato fields. Science of the Total Environment. 958, 177910.
Acknowledgment
Financial support for this project was provided by the Oregon Department of Agriculture — Speciality Crop Block Grant.
References
Ait, K.A.N., B. Galaup, J. Desplanques, G. Dechamp-Guillaume and C. Seassau. (2022). Ecosystem Services Provided by Cover Crops and Biofumigation in Sunflower Cultivation. Agronomy 12, 120.
Ashiq, S., S. Edwards, A. Watson, E. Blundell and M. Back. (2022). Antifungal effect of brassica tissues on the mycotoxigenic cereal pathogen Fusarium graminearum. Antibiotics 11, 1249.
Bajagain, A., E.A. Lehnhoff, R. Creamer, R. Steiner and B.J. Schutte. (2024). Timing Termination of a Biofumigant Cover Crop for Weed Suppression in Chile Pepper. HortTechnology 34, 142-152.
Baysal-Gurel, F., P. Liyanapathiranage and K.M. Addesso. (2020). Effect of Brassica crop-based biofumigation on soilborne disease suppression in woody ornamentals. Canadian Journal of Plant Pathology 42, 94-106.
Bhandari, S.R., J. Rhee, C.S. Choi, J.S. Jo, Y.K. Shin and J.G. Lee. (2020). Profiling of individual desulfo-glucosinolate content in cabbage head (Brassica oleracea var. capitata) germplasm. Molecules 25, 1860.
Brennan, R.J.B., S. Glaze-Corcoran, W. Robert and M. Hashemi. (2020). Biofumigation: An alternative strategy for the control of plant parasitic nematodes. Journal of Integrative Agriculture 19, 1680-1690.
Brown, P., and M. Morra. (1997). Control of Soil-Borne Plant Pests Using Glucosinolate-Containing Plants. Advances in Agronomy 61, 167-231.
Cámara-Martos, F., S. Obregón-Cano, O. Mesa-Plata, M.E. Cartea-González and A. de Haro-Bailón. (2021). Quantification and in vitro bioaccessibility of glucosinolates and trace elements in Brassicaceae leafy vegetables. Food Chemistry 339, 127860.
Campbell, J.W., T.A. Rand, N.M. West, A. Morphew, B.L. Allen, J. Jabro, and S.R. Dangi. (2024). Pollinators and Other Beneficial Insects Within Two Brassicaceous Oilseeds and a Cover Crop Mix Under Evaluation as Fallow Surrogates for Dryland Production Systems of the Northern Great Plains. Journal of the Kansas Entomological Society 96, 78-92.
Carlock, L.L., and T.A. Dotson. (2010). Metam-sodium. In Hayes' Handbook of Pesticide Toxicology, pp. 2293-2306. Elsevier.
Castellano-Hinojosa, A., N.S. Boyd and S.L. Strauss. (2022). Impact of fumigants on non-target soil microorganisms: a review. Journal of Hazardous Materials 427, 128149.
Chen, D., B.J. Zebarth, C. Goyer, L.-P. Comeau, K. Nahar and T. Dixon. (2022). Effect of Biofumigation on population densities of Pratylenchus spp. and Verticillium spp. and potato yield in Eastern Canada. American Journal of Potato Research 99, 229-242.
Collins, H., A. Alva, R. Boydston, R Cochran, P. Hamm, A. McGuire and E. Riga. (2006). Soil microbial, fungal, and nematode responses to soil fumigation and cover crops under potato production. Biology and Fertility of Soils 42, 247-257.
Couëdel, A., L. Alletto, H. Tribouillois and E. Justes. (2018). Cover crop crucifer-legume mixtures provide effective nitrate catch crop and nitrogen green manure ecosystem services. Agriculture, Ecosystems & Environment 254, 50-59.
Dunne, C. (2002). Comparison of integrated management practices for the control of Phytophthora cinnamomi in protea cultivation in WA. Floriculture News.
Edwards, S. and A. Ploeg. (2014). Evaluation of 31 potential biofumigant brassicaceous plants as hosts for three meloiodogyne species. Journal of Nematology, 287-295.
Environmental Protection Agency (2023). Soil Fumigants. Vol. 2023.
Food and Agriculture Organization of the United Nations (2022). Pesticides Trade and Pesticides Indicators — Global, Regional and Country Trends, 1990–2020. FAOSTAT Analytical Briefs 46.
Gimsing, A.L. and J.A. Kirkegaard. (2009). Glucosinolates and biofumigation: fate of glucosinolates and their hydrolysis products in soil. Phytochemistry Reviews 8, 299-310.
Hanschen, F.S., B. Yim, T. Winkelmann, K. Smalla and M. Schreiner. (2015). Degradation of biofumigant isothiocyanates and allyl glucosinolate in soil and their effects on the microbial community composition. PloS one 10, e0132931.
Haramoto, E.R. and E.R. Gallandt. (2004). Brassica cover cropping for weed management: A review. Renewable agriculture and food systems 19, 187-198.
Huang, H., J. Wang, S. Mao, Q. Wu, Y. Tian, F. Wang, P. Wang, K. Huang, and Q. Wu. (2022). Variation characteristics of glucosinolate contents in leaf mustard (Brassica juncea). Agronomy 12, 2287.
Jhingan, S., H.-J. Harloff, A., Abbadi, C. Welsch, M. Blümel, D. Tasdemir and C. Jung. (2023). Reduced glucosinolate content in oilseed rape (Brassica napus L.) by random mutagenesis of BnMYB28 and BnCYP79F1 genes. Scientific Reports 13, 2344.
Kim, J.A., H. Moon, H.S. Kim, D. Choi, N.-S. Kim, J. Jang, S.W. Lee, A. Baskoro Dwi Nugroho and D.-H. Kim. (2023). Transcriptome and QTL mapping analyses of major QTL genes controlling glucosinolate contents in vegetable-and oilseed-type Brassica rapa plants. Frontiers in Plant Science 13, 1067508.
Kim, S.-J., and G. Ishii. (2006). Glucosinolate profiles in the seeds, leaves and roots of rocket salad (Eruca sativa Mill.) and anti-oxidative activities of intact plant powder and purified 4-methoxyglucobrassicin. Soil Science and Plant Nutrition 52, 394-400.
Kirkegaard, J., and J. Matthiessen. (2004). Developing and refining the biofumigation concept. Agroindustria 3, 233-239.
Kruger, D., J. Fourie, and A.P. Malan. (2013). Cover crops with biofumigation properties for the suppression of plant-parasitic nematodes: a review. South African Journal of Enology and Viticulture, 34(2):287
Kumar, V., D.C. Brainard and R.R. Bellinder. (2009). Effects of Spring-sown Cover Crops on Establishment and Growth of Hairy Galinsoga (Galinsoga ciliata) and Four Vegetable Crops. HortScience 44, 730-736.
Larkin, R.P. and C.W. Honeycutt. (2006). Effects of different 3-year cropping systems on soil microbial communities and Rhizoctonia diseases of potato. Phytopathology 96, 68-79.
Lefebvre, M., M. Leblanc and A. Watson. (2019). Impact of Indian mustard growth and incorporation on annual weed population dynamics and communities. Weed Research 59, 324-338.
Li, Y., Y. Yu, L. Xu, E. Guo, Y. Zang, Y. He and Z. Zhu. (2021). Transcriptome profiling reveals candidate key genes involved in sinigrin biosynthesis in Brassica nigra. Horticulturae 7, 173.
Matthiessen, J.N. and J.A. Kirkegaard. (2006). Biofumigation and enhanced biodegradation: opportunity and challenge in soilborne pest and disease management. Critical Reviews in Plant Sciences 25, 235-265.
McGrath, M., and S. Menasha, S. (2013). Managing Phytophthora blight with biofumigation. In Phytopathology, vol. 103, pp. 93-93. Amer Phytopathological Society.
McLeod, R. and C. Steel. (1999). Effects of brassica-leaf green manures and crops on activity and reproduction of Meloidogyne javanica. Nematology 1, 613-624.
Monfort, W., A. Csinos, A., J. Desaeger, K. Seebold, T. Webster,and J. Diaz-Perez, (2007). Evaluating Brassica species as an alternative control measure for root-knot nematode (M. incognita) in Georgia vegetable plasticulture. Crop Protection 26, 1359-1368.
Morra, M., and J. Kirkegaard, J. (2002). Isothiocyanate release from soil-incorporated Brassica tissues. Soil Biology and Biochemistry 34, 1683-1690.
Motisi, N., F. Montfort, V. Faloya, P. Lucas and T. Doré. (2009). Growing Brassica juncea as a cover crop, then incorporating its residues provide complementary control of Rhizoctonia root rot of sugar beet. Field Crops Research 113, 238-245.
Murray, K., P.C. Jepson, I. Sandlin and A. Jensen. (2020). Integrated pest Management strategic plan for potatoes in Oregon, Washington and Idaho, EM 9275.
Murray, K., Reitz, S. R., and Jepson, P. C. (2018). An integrated pest management strategic plan for Treasure Valley onions: Oregon and Idaho, EM 9254.
Murray, K., I. Sandlin, P. Ellsworth, P. Jepson, A. Fournier, H. Luh and S.Reitz. (2022a). The economic impact of onion pests in the Treasure Valley, EM 9347.
Murray, K., I. Sandlin, P. Ellsworth, P.C. Jepson, A. Fournier, H. Luh and S.R. Reitz. (2022b). The economic impact of onion pests in the Treasure Valley: A Look at pests and associated management practices, 2018-2019, Oregon State University Extension Service.
Pattison, A.B., C. Versteeg, S. Akiew and J. Kirkegaard. (2006). Resistance of Brassicaceae plants to root-knot nematode (Meloidogyne spp.) in northern Australia. International Journal of Pest Management 52, 53-62.
Petersen, A., C. Wang, C. Crocoll and B.A. Halkier. (2018). Biotechnological approaches in glucosinolate production. Journal of integrative plant biology 60, 1231-1248.
Roubtsova, T., J.A. López-Péŕez, S. Edwards and A. Ploeg, A. (2007). Effect of Broccoli (Brassica oleracea) Tissue, Incorporated at Different Depths in a Soil Column, on Meloidogyne incognita. Journal of Nematology 39, 111-7.
Rudolph, R. and E. Pfeufer, E. (2021). The Basics of Biofumigation. University of Kentucky College of Agriculture and F. a. Environment, eds.), Vol. CCD-FS-20, Lexington, Kentucky.
Rudolph, R.E., C. Sams, R. Steiner, S.H. Thomas, S. Walker and M.E. Uchanski. (2015). Biofumigation performance of four Brassica crops in a green chile pepper (Capsicum annuum) rotation system in southern New Mexico. HortScience 50, 247-253.
Sennett, L., D.L. Burton, C. Goyer and B.J. Zebarth. (2021). Influence of chemical fumigation and biofumigation on soil nitrogen cycling processes and nitrifier and denitrifier abundance. Soil Biology and Biochemistry 162, 108421.
Smolinska, U., M. Morra, G. Knudsen and R. James. (2003). Isothiocyanates produced by Brassicaceae species as inhibitors of Fusarium oxysporum. Plant Disease 87, 407-412.
Sowmya, R. (2023). Biofumigation for nematode management: Advantages and limitations. Journal of Entomology and Zoology Studies. 11(6): 60-65.
Tsror, L., E. Shlevin and I. Peretz-Alon. (2005). Efficacy of metam sodium for controlling Verticillium dahliae prior to potato production in sandy soils. American Journal of Potato Research 82, 419-423.
Tsytsiura, Y. (2024). Potential of oilseed radish (Raphanus sativus l. var. oleiformis Pers.) as a multi-service cover crop (MSCC). Agronomy Research. 22, No. 2.
Valdes, Y., N. Viaene and M. Moens, M. (2012). Effects of yellow mustard amendments on the soil nematode community in a potato field with focus on Globodera rostochiensis. Applied Soil Ecology 59, 39-47.
Vera, C., D. McGregor and R. Downey, R. (1987). Detrimental effects of volunteer Brassica on production of certain cereal and oilseed crops. Canadian Journal of Plant Science 67, 983-995.
Waddington, J. (1978). Growth of barley, bromegrass and alfalfa in the greenhouse in soil containing rapeseed and wheat residues. Canadian Journal of Plant Science 58, 241-248.
Waisen, P., Z. Cheng, B.S. Sipes, J. DeFrank, S.P. Marahatta and K.-H. Wang. (2020). Effects of biofumigant crop termination methods on suppression of plant-parasitic nematodes. Applied Soil Ecology 154, 103595.
Walker, B.A., S.M. Powell, R.S. Tegg, R.B. Doyle, I.G. Hunt and C.R. Wilson. (2022). Soil microbial community dynamics during ryegrass green manuring and brassica biofumigation. Applied Soil Ecology 179, 104600.
Yan, D., Q. Wang, L. Mao, W. Li, H. Xie, M. Guo and A. Cao. (2013). Quantification of the effects of various soil fumigation treatments on nitrogen mineralization and nitrification in laboratory incubation and field studies. Chemosphere 90, 1210-1215.
Yan, D., Q. Wang, L. Mao, T. Ma, Y. Li, C. Ouyang, M. Guo and A. Cao, A. (2015). Interaction between nitrification, denitrification and nitrous oxide production in fumigated soils. Atmospheric environment 103, 82-86.
Zheng, W., S.R. Yates, S.K. Papiernik, M. Guo and J. Gan. (2006). Dechlorination of chloropicrin and 1, 3-dichloropropene by hydrogen sulfide species: redox and nucleophilic substitution reactions. Journal of Agricultural and Food Chemistry 54, 2280-2287.
About the authors

